§13. Ainevahetus ja energia muundamine rakus. Ainevahetus ja energia rakus Tõestage, et ainevahetus ja energia muundamine
Kõik elusorganismid Maal on avatud süsteemid, mis on võimelised aktiivselt korraldama energia ja aine tarnimist väljastpoolt. Energia on vajalik elutähtsate protsesside läbiviimiseks, aga eelkõige rakkude ja keha struktuuride ehitamiseks ja taastamiseks kasutatavate ainete keemiliseks sünteesiks. Elusolendid on võimelised kasutama ainult kahte tüüpi energiat: valgus(päikese kiirgusenergia) ja keemiline(keemiliste ühendite sidemete energia) - selle alusel jagatakse organismid kahte rühma - fototroofid ja kemotroofid.
Peamine struktuursete molekulide allikas on süsinik. Sõltuvalt süsinikuallikatest jagunevad elusorganismid kahte rühma: autotroofid, mis kasutavad anorgaanilist süsinikuallikat (süsinikdioksiidi) ja heterotroofid, mis kasutavad orgaanilist süsinikuallikat.
Energia ja aine tarbimise protsessi nimetatakse toit. Tuntud on kaks toitumismeetodit: holosoiline – toiduosakeste püüdmise kaudu keha sees ja holofüütne – ilma püüdmiseta, lahustunud toitainete imendumise kaudu läbi keha pinnastruktuuride. Toitained, mis sisenevad kehasse, osalevad ainevahetusprotsessides.
Ainevahetus on omavahel seotud ja tasakaalustatud protsesside kogum, mis hõlmab mitmesuguseid keemilisi muutusi kehas. Energiatarbimisega läbiviidavad sünteesireaktsioonid on anabolismi (plastilise metabolismi või assimilatsiooni) aluseks.
Aluse moodustavad lõhenemisreaktsioonid, millega kaasneb energia vabanemine katabolism(energiavahetus või dissimilatsioon).
1. ATP tähtsus ainevahetuses
Orgaaniliste ainete lagunemisel vabanevat energiat ei kasutata koheselt ära, vaid see salvestub kõrge energiaga ühendite kujul, tavaliselt adenosiintrifosfaadi (ATP) kujul. Oma keemilise olemuse järgi on ATP mononukleotiid ja koosneb lämmastikalusest adeniinist, süsivesikute riboosist ja kolmest fosforhappe jäägist.
ATP hüdrolüüsi käigus vabanevat energiat kasutab rakk igat tüüpi tööde tegemiseks. Märkimisväärne hulk energiat kulutatakse bioloogilisele sünteesile. ATP on universaalne rakuenergia allikas. ATP varu rakus on piiratud ja seda täiendatakse fosforüülimise protsessi tõttu, mis toimub hingamise, fermentatsiooni ja fotosünteesi ajal erineva kiirusega. ATP uueneb ülikiiresti (inimesel on ühe ATP molekuli eluiga alla 1 minuti).
2. Energia metabolism rakus. ATP süntees
ATP süntees toimub kõigi organismide rakkudes fosforüülimise protsessi käigus, s.o. anorgaanilise fosfaadi lisamine ADP-le. ADP fosforüülimiseks vajalik energia tekib energia metabolismi käigus. Energia metabolism ehk dissimilatsioon on orgaaniliste ainete lagunemise reaktsioonide kogum, millega kaasneb energia vabanemine. Olenevalt elupaigast võib dissimilatsioon toimuda kahes või kolmes etapis.
Enamikus elusorganismides - hapnikukeskkonnas elavates aeroobides - viiakse dissimilatsiooni käigus läbi kolm etappi: ettevalmistav, hapnikuvaba, hapnik. Anaeroobides, kes elavad hapnikuvaeses keskkonnas või hapnikuvaegusega aeroobides, toimub dissimilatsioon ainult kahes esimeses etapis, mille käigus moodustuvad vahepealsed orgaanilised ühendid, mis on endiselt energiarikkad.
Esimene etapp - ettevalmistav - koosneb keeruliste orgaaniliste ühendite ensümaatilisest lagundamisest lihtsamateks (valgud aminohapeteks; polüsahhariidid monosahhariidideks; nukleiinhapped nukleotiidideks). Orgaaniliste ainete rakusisene lagunemine toimub lüsosoomide hüdrolüütiliste ensüümide toimel. Sel juhul vabanev energia hajub soojuse kujul ja tekkivad väikesed orgaanilised molekulid võivad läbida täiendava lagunemise ja raku poolt kasutada neid oma orgaaniliste ühendite sünteesiks "ehitusmaterjalina".
Teine etapp - mittetäielik oksüdatsioon - toimub otse raku tsütoplasmas, ei vaja hapniku olemasolu ja koosneb orgaaniliste substraatide edasisest lagunemisest. Raku peamine energiaallikas on glükoos. Glükoosi hapnikuvaba, mittetäielikku lagunemist nimetatakse glükolüüsiks.
Kolmas etapp - täielik oksüdatsioon - toimub hapniku kohustusliku osalusel. Selle tulemusena laguneb glükoosimolekul anorgaaniliseks süsinikdioksiidiks ja sel juhul vabanev energia kulub osaliselt ATP sünteesiks.
3. Plastivahetus
Plastiline ainevahetus ehk assimilatsioon on reaktsioonide kogum, mis tagab keeruliste orgaaniliste ühendite sünteesi rakus. Heterotroofsed organismid loovad oma orgaanilise aine mahetoidu komponentidest. Heterotroofne assimilatsioon taandub sisuliselt molekulide ümberpaigutamisele.
Toidu orgaanilised ained (valgud, rasvad, süsivesikud) --> seedimine --> lihtsad orgaanilised molekulid (aminohapped, rasvhapped, monosahhariidid) --> bioloogiline süntees --> keha makromolekulid (valgud, rasvad, süsivesikud)
Autotroofsed organismid on võimelised väliskeskkonnast tarbitavatest anorgaanilistest molekulidest täiesti sõltumatult sünteesima orgaanilisi aineid. Autotroofse assimilatsiooni protsessis eelnevad makromolekulide bioloogilisele sünteesile foto- ja kemosünteesi reaktsioonid, mis tagavad lihtsate orgaaniliste ühendite moodustumise:
Anorgaanilised ained (süsinikdioksiid, vesi) --> fotosüntees, kemosüntees --> lihtsad orgaanilised molekulid (aminohapped, rasvhapped, monosahhariidid) -----bioloogiline süntees --> keha makromolekulid (valgud, rasvad, süsivesikud)
4. Fotosüntees
Fotosüntees on orgaaniliste ühendite süntees anorgaanilistest ühenditest, kasutades raku energiat. Fotosünteesi protsessides mängivad juhtivat rolli fotosünteesi pigmendid, millel on ainulaadne omadus püüda valgust ja muuta selle energia keemiliseks energiaks. Fotosünteetilised pigmendid on üsna suur rühm valgutaolisi aineid. Peamine ja kõige olulisem energeetiliselt on pigment. klorofüll a, mida leidub kõigis fototroofides, välja arvatud fotosünteetilistes bakterites. Fotosünteetilised pigmendid on põimitud eukarüootidel plastiidide sisemembraani või prokarüootidel tsütoplasmaatilise membraani sissetungidesse.
Fotosünteesi käigus sünteesitakse lisaks monosahhariididele (glükoos jne), mis muundatakse tärkliseks ja säilitatakse taime poolt, teiste orgaaniliste ühendite monomeerid - aminohapped, glütserool ja rasvhapped. Seega tagavad taimerakud, täpsemalt klorofülli sisaldavad rakud, tänu fotosünteesile endale ja kogu Maal elavale olendile vajalike orgaaniliste ainete ja hapnikuga.
5. Kemosüntees
Kemosüntees on ka orgaaniliste ühendite sünteesimise protsess anorgaanilistest, kuid see ei toimu mitte valgusenergia, vaid anorgaaniliste ainete (väävel, vesiniksulfiid, raud, ammoniaak, nitrit jne). Olulisemad on nitrifitseerivad, raua- ja väävlibakterid.
Oksüdatsioonireaktsioonide käigus vabanev energia salvestatakse bakterite poolt ATP kujul ja seda kasutatakse orgaaniliste ühendite sünteesiks. Kemosünteetilistel bakteritel on biosfääris väga oluline roll. Nad osalevad reoveepuhastuses, aitavad kaasa mineraalide kogunemisele pinnasesse ja suurendavad mulla viljakust.
DNA - biopolümeer, mikromolekul, polünukleotiid, -manomeer-nukleotiid Lämmastiku alused - desoksüriboos - fosforhappe jääk Lämmastiku alused: adeniin, tümiin, guaniin, tsütosiin - RNA kaheahelaline struktuur - biopolümeer, makromolekul, nikotiini polünukleotiid - Riboos - Fosforhappe jääk Lämmastiku alused: adeniin, uratsiil, guaniin, tsütosiin. RNA molekul on üheahelaline. Funktsioonid: DNA – geneetilise informatsiooni talletamine RNA – geneetilise informatsiooni edastamine
Tuumas sünteesitakse Messenger RNA, mis kannab infot valgumolekulide esmase struktuuri kohta. Pärast tuumamembraani pooride läbimist saadetakse mRNA ribosoomidesse, kus geneetiline informatsioon dešifreeritakse - tõlgitakse nukleotiidide keelest aminohapete keelde.
Aminohapped, millest valke sünteesitakse, viiakse ribosoomidesse spetsiaalsete RNA-de abil, mida nimetatakse ülekande-RNA-deks (t-RNA). t-RNA-s on kolme nukleotiidi järjestus komplementaarne i-RNA koodoni nukleotiididega. Seda nukleotiidide järjestust tRNA struktuuris nimetatakse antikoodoniks. Iga t-RNA seob ensüüme ja ATP-d kasutades spetsiifilise aminohappe. See on sünteesi esimene etapp.
Selleks, et aminohape kuuluks valguahelasse, peab see tRNA-st lahti murdma. Valgusünteesi teises etapis toimib tRNA tõlkijana nukleotiidide keelest aminohapete keelde. See translatsioon toimub ribosoomil. Selles on kaks sektsiooni: ühel saab t-RNA käsu mRNA-lt - antikoodon tunneb koodoni ära, teiselt poolt täidetakse käsk - aminohape rebitakse t-RNA-lt ära.
Valgusünteesi kolmas etapp seisneb selles, et ensüümi süntetaas seob tRNA-st eraldunud aminohappe kasvava valgu molekuliga. Messenger RNA libiseb pidevalt mööda ribosoomi, iga kolmik langeb esmalt esimesse sektsiooni, kus ta tunneb ära tRNA antikoodon, seejärel teise sektsiooni. Siia läheb ka t-RNA koos selle külge kinnitatud aminohappega, siin eraldatakse aminohapped t-RNA-st ja ühendatakse üksteisega järjestuses, milles kolmikud järgnevad üksteise järel.
Kui esimeses osas ilmub ribosoomile üks kolmest kolmikust, mis on geenidevahelised kirjavahemärgid, tähendab see, et valgusüntees on lõppenud. Valmis valguahel lahkub ribosoomist. Valgu sünteesi protsess nõuab palju energiat. Iga aminohappe ühendamiseks t-RNA-ga on vaja ühe ATP molekuli energiat.
Valgu tootmise suurendamiseks läbib mRNA sageli samaaegselt mitte ühte, vaid mitut ribosoomi järjest. Sellist struktuuri, mida ühendab üks mRNA molekul, nimetatakse polüsoomiks. Igal ribosoomil, helmeste stringiga sarnasel konveieril sünteesitakse järjestikku mitu identsete valkude molekuli.
Valkude sünteesi ribosoomidel nimetatakse translatsiooniks. Valgumolekulide süntees toimub pidevalt ja toimub suurel kiirusel: ühe minuti jooksul moodustub 50 kuni 60 tuhat peptiidsidet. Ühe valgumolekuli süntees kestab vaid 3-4 sekundit. Iga biosünteesi etappi katalüüsivad sobivad ensüümid ja ATP lagunemise kaudu varustatakse neid energiaga. Sünteesitud valgud sisenevad endoplasmaatilise retikulumi kanalitesse, mille kaudu transporditakse need raku teatud osadesse.
Taimerakk kui osmootne süsteem
Taimerakk on osmootne süsteem. Vakuooli rakumahl on väga kontsentreeritud lahus. Rakumahla osmootne rõhk on tähistatud -.
Vakuooli sisenemiseks peab vesi läbima rakuseina, plasmalemma, tsütoplasma ja tonoplasti. Rakusein on vett hästi läbilaskev. Plasmalemmal ja tonoplastil on selektiivne läbilaskvus. Seetõttu võib taimerakku käsitleda osmootse süsteemina, milles plasmamembraan ja tonoplast on poolläbilaskev membraan ning rakumahlaga vakuool on kontsentreeritud lahus. Seega, kui rakk asetatakse vette, hakkab vesi vastavalt osmoosi seadustele rakku voolama.
Jõudu, millega vesi rakku siseneb, nimetatakse imemisjõuks - S.
See on identne veepotentsiaaliga.
Kui vesi siseneb vakuooli, suureneb selle maht, vesi lahjendab rakumahla ja rakuseinad hakkavad kogema survet. Rakusein on teatud elastsusega ja võib venida.
Kui vakuooli maht suureneb, surutakse tsütoplasma vastu rakuseina ja rakuseinale tekib turgorurve (P). Samal ajal tekib rakuseinast võrdne vasturõhk protoplastile. Rakuseina vasturõhku nimetatakse survepotentsiaaliks (-P).
Seega määrab imemisjõu S suuruse rakumahla osmootne rõhk ja raku P turgori hüdrostaatiline rõhk, mis on võrdne rakuseina vasturõhuga, mis tekib selle venitamisel -P.
S = - P või - - .
Kui taim on piisava mulla- ja õhuniiskuse tingimustes, siis on rakud täielikus turgoris. Kui rakk on veega täielikult küllastunud (turgesents), on selle imemisjõud null S = 0 ja turgori rõhk võrdub potentsiaalse osmootse rõhuga P =.
Kui mullas on niiskusepuudus, tekib veepuudus esmalt rakuseinas. Rakuseina veepotentsiaal muutub madalamaks kui vakuoolis ja vesi hakkab liikuma vakuoolist rakuseinasse. Vee väljavool vakuoolist vähendab turgorirõhku rakkudes ja suurendab nende imemisjõudu. Pikaajalise niiskuse puudumise korral kaotab enamik rakke turgori ja taim hakkab närbuma, kaotades elastsuse ja tugevuse. Sel juhul turgorirõhk P = 0 ja imemisjõud S =
Kui väga suure veekao tõttu langeb turgori rõhk nullini, närbub leht täielikult. Edasine veekadu põhjustab raku protoplasti surma. Kohanemisomadus äkilise veekaotusega on stoomide kiire sulgumine niiskusepuuduse korral.
Rakud suudavad turgori kiiresti taastada, kui taim saab piisavalt vett või öösel, kui taim saab mullast piisavalt vett. Ja ka kastmisel.
Vee potentsiaal; võrdub 0 puhta vee puhul; võrdub 0-ga või rakkude puhul negatiivne.
Osmootne potentsiaal on alati negatiivne
Survepotentsiaal; tavaliselt positiivne elusrakkudes (rakkudes, mille sisu on rõhu all), kuid negatiivne ksüleemrakkudes (milles tekib vee pinge).
Tegevuse kogutulemus
Täieliku turgoriga
Esialgse plasmolüüsi ajal
Kui asetada rakk madalama veepotentsiaaliga hüpertoonilisse lahusesse, hakkab vesi rakust väljuma osmoosi teel läbi plasmamembraani. Esiteks lahkub vesi tsütoplasmast, seejärel läbi tonoplasti vakuoolist. Raku elussisu, protoplast, kahaneb ja langeb rakuseina taha. Toimub protsess plasmolüüs. Rakuseina ja protoplasti vaheline ruum täidetakse välise lahusega. Sellist rakku nimetatakse plasmolüüsitud rakuks. Vesi lahkub rakust, kuni protoplasti veepotentsiaal muutub võrdseks ümbritseva lahuse veepotentsiaaliga, misjärel rakk kahanemise lõpetab. See protsess on pöörduv ja rakk ei kahjusta.
Kui rakk asetada puhtasse vette või hüpotoonilisse lahusesse, taastub raku turgori olek ja protsess deplasmolüüs.
Noorte kudede veepuuduse tingimustes viib veekao järsk suurenemine selleni, et raku turgorirõhk muutub negatiivseks ja mahult kokkutõmbuv protoplast ei eraldu rakuseinast, vaid tõmbab selle endaga kaasa. . Rakud ja kuded kahanevad. Seda nähtust nimetatakse tsütorhiis.
Ainete ja energia ainevahetus (ainevahetus) toimub keha kõigil tasanditel: raku-, koe- ja organismitasanditel. See tagab keha sisekeskkonna – homöostaasi – püsivuse pidevalt muutuvates eksistentsitingimustes. Rakus toimuvad samaaegselt kaks protsessi – plastiline ainevahetus (anabolism või assimilatsioon) ja energia metabolism (fatabolism või dissimilatsioon).
Plastiline vahetus on kõigi sünteesiprotsesside kogum, kui lihtsatest ainetest moodustuvad keerukad ained, samal ajal kui energiat kulutatakse.
Energiaainevahetus on kõigi lõhustamisprotsesside kogum, kui keerulised ained moodustuvad lihtsateks ja vabaneb energia.
Homöostaasi säilitab tasakaal plasti ja energia metabolismi vahel. Kui see tasakaal on häiritud, tekivad kehas või selle osas patoloogiad (haigused).
Ainevahetus toimub normaalsel temperatuuril, rõhul ja teatud pH keskkonnas
11.Energia metabolism rakus.
Energia metabolism on orgaaniliste ühendite järkjärgulise lagunemise keemiliste reaktsioonide kogum, millega kaasneb energia vabanemine, millest osa kulub ATP sünteesiks. Sünteesitud ATP-st saab universaalne energiaallikas organismide eluks.
Energia metabolismi etapid:
1. Ettevalmistav - sellel jagatakse keerulised ained lihtsateks, näiteks polüsahhariidid monosahhariidideks. See etapp toimub tsütoplasmas ja vabastab energiat, kuid seetõttu hajub soojusena väga vähe energiat.
2. Hapnikuvaba - lüsosoomides jätkub selles etapis ainete lagunemine lihtsamateks ilma hapniku osaluseta kahe ATP molekuli vabanemisega
3. Hapnik - see jätkab ainete lagunemist hapniku osalusel lõpptoodeteks (süsinikdioksiid ja vesi) koos 36 ATP vabanemisega. See protsess toimub mitokondrites.
Rakkude toitumine. Kemosüntees
Rakkude toitumine toimub mitmete keerukate keemiliste reaktsioonide tulemusena, mille käigus väliskeskkonnast rakku sisenevad ained (süsinikdioksiid, mineraalsoolad, vesi) sisenevad raku enda kehasse valkude, suhkrute, rasvade kujul. , õlid, lämmastik ja fosfor.ühendused.
Kõik elusorganismid võib jagada kahte rühma:
1. Autotroofne toitumistüüp – nende hulka kuuluvad organismid, kes ise sünteesivad orgaanilisi ühendeid anorgaanilistest.
2 tüüpi autotroofe:
Fotosünteetilised ained on autotroofid, mis kasutavad päikesevalguse energiat (taimed, tsüanobakterid, algloomad)
Kemosünteetikumid on organismid, mis kasutavad keemiliste sidemete energiat. Sellesse tüüpi kuuluvad peaaegu kõik bakterid (lämmastikufiksaatorid, väävlibakterid, rauabakterid)
Kemosünteesi avastas Vinogradov.
Kemosüntees on autotroofse toitumise meetod, mille puhul CO2-st orgaaniliste ainete sünteesi energiaallikaks on anorgaaniliste ühendite oksüdatsioonireaktsioonid. Seda energia saamise võimalust kasutavad ainult bakterid või arheed.
2. Heterotroofne toitumistüüp – omane organismidele, kes toituvad valmisorgaanilistest ühenditest.
Soprofüüdid on heterotroofid, mis toituvad surnud kudedest või organismidest (varesed, raisakotkad, hüäänid...)
Taimtoidulised - heterotroofid, kes söövad taimseid organisme (rohutoidulised)
Kiskjad (kiskjad) on heterotroofid, kes püüavad ja söövad teisi organisme (putuktoidulised)
Kõigesööjad – söövad taimset ja loomset toitu
3. Miksotroofne toitumistüüp - ühendab autotroofseid ja heterotroofseid toitumistüüpe (sundew, roheline euglena)
Fotosüntees
Fotosüntees on keeruline protsess anorgaaniliste ainete moodustumiseks päikesevalguse energia abil. Peamine fotosünteesi organ on leht, kuna see sisaldab kõige rohkem kloroplaste ja selle kuju on päikesevalguse vastuvõtmiseks kõige sobivam.
Fotosünteesi faasid:
1. Kerge faas – sisaldab 2 põhiprotsessi: vee fotolüüs ja mittetsükliline fosforüülimine.
Tülakoidid on lamestatud membraanikotid, millel paiknevad klorofüllipigmendid ja spetsiaalne elektronide kandja, mida nimetatakse tsütokroomiks.
Tülakoididel on 2 fotosüsteemi:
Fotosüsteem 1 sisaldab klorofülli a1, mis tajub valguskvanti pikkusega 700 nanomeetrit
Photosystem 2 sisaldab klorofülli a2, mis tajub valguskvanti pikkusega 680 nanomeetrit
Kui valguskvant tabab fotosüsteemi 1, ergastuvad klorofülli a1 elektronid ja kantakse üle sellisesse protsessi nagu vee fatolüüs, st vesi jaguneb vesinikuks ja hüdroksorühmaks. Aine redutseerimiseks kasutatakse vesinikku. Saadud hüdroksorühm koguneb ja muundatakse veeks ja hapnikuks, mis lahkub rakust.
Kui valguskvant tabab fotosüsteemi 2, ergastuvad valguse mõjul klorofülli elektronid ja ADP molekulile lisandub energia toimel fosforhappejääk, mille tulemusena tekib ATP molekul.
Valgusfaas toimub tülakoodidel, kus tekib orgaaniliste ainete tekkeks vajalik energia.
Tume faas – esineb stroomas päikesevalgusest sõltumatult. Siin muudetakse keeruliste reaktsioonide käigus tekkinud energia abil süsihappegaas glükoosiks. Neid reaktsioone nimetatakse Calvini tsükliks.
Geneetiline kood
See on kõigile elusorganismidele omane meetod valkude aminohappejärjestuse kodeerimiseks nukleotiidide järjestuse abil
DNA võib sisaldada 4 lämmastiku alust:
Adeniin, guaniin, tümiin, tsütosiin
DNA suudab kodeerida 64 aminohapet
Omadused:
1. Degeneratsioon – suurendab rakkude jagunemise ajal geneetilise informatsiooni säilitamise ja edastamise usaldusväärsust
2. Spetsiifilisus – 1 triplett kodeerib alati ainult 1 aminohapet
Geneetiline ko on universaalne kõigile elusorganismidele alates bakteritest kuni inimesteni
15. Transkriptsioon ja saade
Valkude süntees koosneb kahest etapist:
1. Transkriptsioon on informatsiooni transkriptsioon DNA molekulist messenger RNA-ks
See protsess toimub tuumas ensüümi RNA polümeraasi osalusel. See ensüüm määrab sünteesi alguse ja lõpu. Algus on spetsiifiline nukleotiidide järjestus, mida nimetatakse promootoriks. Lõpp on ka nukleotiidide jada, mida nimetatakse terminaatoriks.
Transkriptsioon algab DNA molekuli lõigu määramisega, millest teave kopeeritakse
Seejärel keritakse see osa lahti vastavalt ühe DNA ahela komplementaarsuse põhimõttele ja luuakse messenger RNA. Kui DNA süntees on lõppenud, keerdub see uuesti.
2. Translatsioon on messenger-RNA nukleotiidjärjestuse translatsioon aminohappejärjestuseks
Transfer RNA kannab messenger RNA ribosoomi. Siin on messenger RNA integreeritud ribosoomi väikesesse subühikusse, kuid sinna mahub ainult 2 kolmikut, mistõttu sünteesi käigus liigub messenger RNA suurde subühikusse, ülekande RNA kannab aminohappeid, kui aminohape sobib, siis on eraldatakse ülekande-RNA-st ja kinnituvad peptiidipõhiste ühenduste kohaselt teiste aminohapetega.
Transfer RNA lahkub ribosoomist ja uued ülekande-RNA-d sisenevad suurde subühikusse
Kui aminohape ei ühti komplementaarsuse põhimõtte kohaselt väikeses subühikus oleva informatsiooniga, siis see transpordi-RNA koos aminohappega lahkub ribosoomist.
Valgusünteesi algust näitavad adeniin, uratsiil, guaniin ja lõpeb stop-kadooniga
Kui valgusüntees lõpeb, eraldatakse valgu esmane struktuur ribosoomist ja valk omandab soovitud struktuuri
Raku elutsükkel
Rakutsükkel on raku eksisteerimise periood alates selle moodustumise hetkest emaraku jagunemise teel kuni tema enda jagunemiseni või surmani.
Interfaas on elutsükli faas kahe raku jagunemise vahel. Seda iseloomustavad aktiivsed ainevahetusprotsessid, valkude ja RNA süntees, toitainete kogunemine raku poolt, kasv ja mahu suurenemine. Interfaasi keskel toimub DNA dubleerimine (replikatsioon). Selle tulemusena sisaldab iga kromosoom 2 DNA molekuli ja koosneb kahest sõsarkromatiidist, mis on omavahel seotud tsentromeeriga ja moodustavad ühe kromosoomi. Rakk valmistub jagunemiseks, kõik selle organellid kahekordistuvad. Interfaasi kestus sõltub raku tüübist ja moodustab keskmiselt 4/5 raku elutsükli koguajast. Raku pooldumine. Organismi kasv toimub tema rakkude jagunemise kaudu. Jagunemisvõime on raku elu kõige olulisem omadus. Kui rakk jaguneb, kahekordistab see kõik oma struktuurikomponendid, mille tulemuseks on kaks uut rakku. Levinuim rakkude jagunemise meetod on mitoos – kaudne rakkude jagunemine. Mitoos on protsess, mille käigus toodetakse kaks algse emarakuga identset tütarrakku. See tagab rakkude uuenemise vananemisprotsessi ajal. Mitoos koosneb neljast järjestikusest faasist:
1. Profaas - kahe kromatiidiga kromosoomide moodustumine, tuumamembraani hävitamine.
2.Metofaas – spindli teke, kromosoomide lühenemine, ekvaatoriraku teke
3. Anafaas - kromatiidide eraldamine, nende lahknemine poolustele piki spindli kiude
4. Telofaas – spindli kadumine, tuumamembraanide moodustumine, kromosoomide lagunemine.
Mitoos. Amitoos
Mitoos on eukarüootide somaatiliste rakkude kaudse jagunemise protsess, mille tulemusena pärandmaterjal algul kahekordistub ja seejärel jaotub tütarrakkude vahel ühtlaselt. See on eukarüootsete rakkude peamine jagunemisviis. Mitoosi kestus loomarakkudes on 30-60 minutit ja taimerakkudes 2-3 tundi. See koosneb 4 põhifaasist:
1. Profaas - algab DNA ahelate speraliseerumisega kromosoomideks, tuumad ja tuumamembraan hävivad, kromosoomid hakkavad tsütoplasmas vabalt hõljuma Profaasi lõppedes hakkab tekkima spindel
2. Metafaas – kromosoomid reastuvad rangelt ekvaatoril metafaasiplaadi kujul. Spindli niidid, mis on juba täielikult moodustunud, läbivad kromosoomide tsentromeere, jagades kromosoomi kaheks kromatiidiks
3. Anafaas – siin eralduvad spindli filamendid ja venivad kromatiidi erinevatele poolustele. Lõhustumisspindel hakkab kokku kukkuma.
4. Telofaas Siin, raku poolustel, kromatiidid hajutatakse, kaetakse tuumamembraaniga ning algab tsütoplasma ja raku enda jagunemine.
Mitoosi tulemusena moodustub 2 identset diploidset rakku.
Karüokenees on tuumajaotus
Tsütokenees on tsütoplasma ja raku enda jagunemine
Amitoos on tuuma otsene jagunemine, mille tulemusena moodustub kahe tuumaga rakk, see tüüp on iseloomulik lihasrakkudele ja sidekudedele
See on vajalik raku töö täielikuks korraldamiseks.
Kui selline rakk äkki jaguneb, sisaldavad uued rakud mittetäielikku geneetilist komplekti, mis põhjustab nende surma või muudab need patogeeniks.
Meioos
See on sugurakkude kaudne jagunemine, mille tulemusena moodustub 4 erineva geneetilise materjaliga haploidset tütarrakku. See on sugurakkude moodustumise peamine etapp.
Meioosi bioloogiline tähtsus:
1. Tänu meioosile tekivad geneetiliselt erinevad sugurakud
2. Somaatilistes rakkudes säilib kromosoomide diploidse komplekti püsivus
3. Tänu meioosile toodab 1 rakk 4 uut rakku
Meioos koosneb kahest osast:
Reduktsioon - selle jagunemise käigus kromosoomide arv väheneb
Võrdlus - kulgeb samamoodi nagu mitoos
Interfaas kulgeb samamoodi nagu mitoos, st DNA kahekordistub jaguneva raku tuumas.
1 meiootiline jaotus
Profaas on meioosi kõige keerulisem ja pikim faas, kuna siin ilmub 2 täiendavat protsessi.
1- Konjugatsioon on homoloogsete kromosoomide lähedane lähenemine, mille tulemusena moodustub 4 kromatiidi, mida ühendab 1 tsentromeer ja sellist struktuuri nimetatakse kahevalentseks. Seejärel toimub kahevalentseks ühendatud kromosoomide vahel ristumine.
2- Crossing over - kromosoomiosade vahetus. Nende protsesside tulemusena toimub 1 geeni rekombinatsioon
Metafaas - siin, raku ekvaatoril, moodustavad bivalentsid metafaasiplaadi, mille tsentromeeridest läbivad ka spindli filamendid
Anafaas – erinevalt mitoosist hajuvad siin terved kromosoomid raku poolustele. Siin toimub 2 geenirekombinatsiooni
Telofaas - loomadel ja mõnel taimel hakkavad kromosoomid lahti rulluma, kaetakse pooluste juures tuumamembraaniga ja jagunevad kaheks rakuks (ainult loomadel)
Taimedel tekib pärast anafaasi kohe profaas 2.
Interfaas on iseloomulik ainult loomadele, erinevalt mitoosi interfaasist ei suurene pärilik teave
Meioosi 2. osakonda kuuluvad profaas, metafaas, telofaas, anafaas, mis kulgevad täpselt nagu mitoosis, kuid vähemate kromosoomidega.
Mittesuguline paljunemine.
See on paljunemise tüüp, mida iseloomustavad:
2. Osaleb 1 üksikisik
3. esineb soodsatel tingimustel
4. kõik organismid osutuvad ühesugusteks
5. säilitab stabiilselt muutumatute tingimuste omadused ja omadused
Bioloogiline tähtsus:
1. vajalik identsete anatoomiliste omadustega organismide tekkeks
2. evolutsioonilises mõttes ei ole mittesuguline paljunemine tulus, kuid tänu sellele paljunemisele suureneb isendite arv populatsioonis lühikese ajaga
Mittesugulise paljunemise tüübid:
Mitootiline jagunemine – tekib mitoosi tõttu (amööb, vetikad, bakterid...)
Sporulatsioon toimub eoste, seente ja taimede spetsiaalsete rakkude kaudu. Kui eosel on vibur, siis nimetatakse seda zoospooriks ja see on iseloomulik veekeskkonnale (eosed, seened, samblikud...)
Küürus - emaindindil tekib väljakasv - pung (sisaldab tütartuuma), millest areneb uus isend.Pung kasvab ja saavutab emaisendi suuruse, alles siis eraldub temast (Hydra, pärmseened, imemine ripsloomad)
Vegetatiivne - paljudele taimerühmadele omane, uus isend areneb kas eristruktuuridest või osast emaisendist.
Mõnedel mitmerakulistel loomadel on ka vegetatiivne paljunemine (käsnad, meritähed, lestaussid)
Seksuaalne paljunemine
Iseloomulik:
Osaleb 1,2 organisatsiooni
2. kaasatud on sugurakud
3. lapsed osutuvad mitmekesiseks
4. evolutsioonilises mõttes ilmnes see hiljem kui aseksuaalne
5. esineb ebasoodsates tingimustes
Bioloogiline tähtsus:
1. järglased on paremini kohanenud muutuvate keskkonnatingimustega ja elujõulisemad
2. tekivad uued organismid
Patogenees (neitsisigimine)
Tütarorganismid arenevad viljastamata munadest.
Patogeneesi tähendus:
1. Paljunemine on võimalik eri soost organismide harvaesinevate kontaktidega
2. Vajalik arvukuse maksimeerimiseks kõrge suremusega populatsioonides
3. Mõne populatsiooni arvukuse hooajaliseks kasvuks
1. Kohustuslik (kohustuslik) – leidub populatsioonides, kus on ainult emased isendid (kaukaasia kivisisalik)
2. Tsükliline (hooajaline) - iseloomulik lehetäidele, planktonile, dafniale, leidub populatsioonides, mis teatud aastaajal hüsteeriliselt välja surevad.
3. Fakultatiivne (ei ole kohustuslik) - leidub sotsiaalsetes putukates. Isased väljuvad viljastamata munadest ja töölised putukad viljastatud munadest.
Sugurakkude areng
Gametogenees
Sugurakud on sugurakud, mis ühinevad, moodustades sigooti, millest areneb uus organism.
Somaatiliste rakkude ja sugurakkude erinevus:
1 sugurakud kannavad haploidset kromosoomikomplekti ja somaatilised diploidi
2. sugurakud ei jagune, küll aga somaatilised
3. sugurakud, eriti somaatilistest rakkudest suuremad munad
Gametogenees on sugurakkude teke, mis esinevad sugunäärmetes (munasarjades, munandites)
Oogenees on gametogenees, mis toimub naise kehas ja viib naise sugurakkude (munaraku) moodustumiseni.
Spermatogenees on gametogenees, mis toimub mehe kehas ja viib meessugurakkude (sperma) moodustumiseni.
Gametogenees koosneb mitmest etapist:
1. Paljunemine – siin, esmastest sugurakkudest, mida nimetatakse spermatogooniaks ja oogooniaks, suureneb tulevaste sugurakkude arv mitoosi kaudu. Spermatogooniad paljunevad meessoost kehas kogu paljunemisperioodi jooksul.
Naise kehas esineb 1. staadium 2–5 kuud emakasisesest arengust.
2. Kasv – esmased sugurakud suurenevad ja muutuvad esimest järku munarakkudeks ja spermatotsüütideks. Need rakud moodustuvad interfaasis. Selles etapis algab meioos.
3. Küpsemine – toimub kahes järjestikuses jaotuses – taandamine ja võrrand. Meioosi 1. jagunemise tulemusena moodustuvad teist järku munarakud ja spermatotsüüdid, pärast 2. meioosi jagunemist spermatotsüütidest 4 spermotiidi.
Teist järku munarakkudest moodustub 1 suur munarakk ja 3 redutseerivat keha. See on tingitud sellest, et kogu energia ja toitained lähevad 1 suure suguraku tekkeks ning ülejäänud 3 raku tekkeks ei jätku jõudu.
Seetõttu jagatakse reprodutseerimiskoodis 3 redutseerimiskeha
4. Tekkimine - selles etapis kasvavad, arenevad spermatiidid ehk täielikult moodustunud sugurakud, omandavad lipu ja täiskasvanud suguraku kuju. Spermatiidid toodetakse spermatosoididest.
Spermatosoidid moodustuvad peast, kaelast ja sabast.
Muna on sarnane somaatilise rakuga, ainult et see on suurema suurusega ja sellel on täiendavad membraanid.
Väetamine
See on sugurakkude ühinemise protsess, mille tulemusena moodustub sügoot – see on uue organismi esimene rakk
1. Väline – seda tüüpi viljastamisega lükkab emane mängimise edasi ja isane kastab teda seemnevedelikuga. Seda tüüpi esineb ainult veekeskkonnas. Spetsiaalseid paljunemisstruktuure pole vaja, pärilikkusmaterjali toodetakse suures koguses ja järglaste ellujäämisprotsent on minimaalne.
2. Sisemine - selle tüübi puhul paigutatakse meeste sugurakud naiste suguelunditesse. See tüüp nõuab spetsiaalseid reproduktiivstruktuure. Pärilikkust toodetakse vähem. Järglaste elulemus suureneb. Niipea, kui meessoost sugurakud sisenevad emase reproduktiivtrakti, liiguvad nad sihikindlalt munaraku suunas, kui üks sperma tungib munarakku, muutuvad selle membraanid tihedamaks ja see muutub teistele spermatosoididele kättesaamatuks. See on vajalik organismide diploidsuse säilitamiseks.
Topeltväetamine
Iseloomulik ainult katteseemnetaimedele. Tolmudes jagunevad primaarsed isased sugurakud meioosi teel, moodustades 4 mikrospoori, iga mikrospoor jaguneb jälle 2 rakuks (vegetatiivsed ja generatiivsed)
Need rakud on kaetud topeltmembraaniga, moodustades õietolmu tera
Mesikas moodustub primaarsest emasrakust meioosi teel 1 megaspoor ja 3 rakku sureb. Saadud megaspoor jaguneb endiselt 2 rakuks, 1 on eoses kesksel kohal ja 2 läheb alla
Õietolmu tera maandub emaka stigmale, vegetatiivne rakk idaneb, moodustades munasarjale õietolmutoru. Generatiivne rakk laskub läbi selle toru ja jaguneb kaheks spermaks. 1 sperma viljastab keskrakku, millest moodustub endosperm.
2 spermat viljastavad teist rakku, millest embrüo areneb.
Ontogenees
See on sügoodi (organismi) individuaalne areng kuni tema surmani. Termini kehtestas 1866. aastal Ernest Haeckel
Imetajatel reguleerivad otnogeneesi närvi- ja endokriinsüsteem
1. Vastne - seda tüüpi munakoorest väljudes jääb organism mõnda aega vastse staadiumisse, seejärel läbib metamorfoosi (muutub täiskasvanuks)
2. Oviparous - seda tüüpi arengu korral püsib organism munakestes pikka aega ja vastse staadium puudub
3. Emakasisene - siin toimub keha areng ema keha sees
Ontogeneesi perioodid:
1. Embrüonaalne (emakasisene) viljastumisest sünnini
2. Postembrüonaalne – sünnist surmani
Embrüonaalne periood
3 arenguetappi
1. Purustamine
Algab paar tundi pärast viljastamist. Siin hakkab sügoot mitootiliselt jagunema 2 rakuks (blastomeerid).Need rakud ei lahkne ega kasva. Seejärel jagunevad need rakud uuesti ja moodustavad 4 rakku ning see jätkub kuni moodustub 32 rakku, kuni moodustub morula - see on embrüo, mis koosneb 32 väikesest vaarika meenutavast rakust ja sügoodi suurusest.
See morula laskub mööda munajuha emakaõõnde ja implanteerub selle seina. See juhtub 6 tundi pärast viljastamist.
Seejärel jätkavad morula rakkude jagunemine ja moodustub blastula - see on mitmesajast rakust koosnev embrüo, mis paikneb 1 kihis.Blastula on õõnsusega ja selle suurus on sama kui sügoodil
2. Gastrulatsioon
Sisaldab blastulat ja gastrulat
Blastula jaguneb edasi ja ühes otsas on rakkude jagunemine intensiivsem. See viib nende rakkude tungimiseni blastulasse, st moodustub gastrula
Gastrula on kahekihiline esmase suuga embrüo, mis imetajatel ja kõrgematel organismidel arenedes muutub pärakuks. Ja õige suu moodustub teisest otsast. Gastrula õõnsus on esmane rakk.
Rakkude välimine kiht on ektoderm (1 idukiht)
Rakkude sisemine kiht on endoderm (2 pakendilehte)
Seejärel moodustub esmase suu mõlemas otsas sümmeetriliselt ektodermi ja endodermi vahele 3 idukihti (mesoderm).
3. Organogenees
Selles etapis moodustub neurula, embrüo seljaosas moodustab raku välimine kiht soone, mis sulgub ja moodustab neuraaltoru. Paralleelselt selle protsessiga moodustub endodermist sooletoru. Ja mesodermist moodustub notokord. Ektodermist moodustub närvisüsteem ja meeleelundid, samuti surnuepiteel ja selle derivaadid (juuksed, küüned).
endoderm – moodustab seedesüsteemi ja seedenäärmed, hingamissüsteemi ja kilpnääre.
4. Mesoderm
Moodustuvad luu- ja lihaskonna süsteem, vereringe-, eritus- ja reproduktiivsüsteem.
Postembrüonaalne periood
Postembrüonaalne areng võib toimuda kahel viisil:
Otsene ja kaudne: täieliku ja mittetäieliku teisendusega
Otsene areng on tüüpiline lindudele, kaladele, imetajatele ja inimestele. Uus isend on sündides ja munakoortest väljudes sarnane täiskasvanud isendiga, kuid väikese suurusega, erinevate proportsioonidega, vähearenenud närvi- ja reproduktiivsüsteemiga, samuti võib erineda nahk.
Postembrüonaalse arengu käigus arenevad edasi närvi- ja reproduktiivsüsteem. Kate vahetub ja keha läbib koolituse ja koolituse.
Kaudne areng – selle tüübi puhul esineb postembrüonaalses arengus vastse staadium. Vastne sarnaneb täiskasvanud loomaga vähe või üldse mitte. Ta kasvab intensiivselt, areneb ja sööb palju toitu.
Seda tüüpi kaudse arengu korral läbib munast väljuv organism vastse staadiumi, mis muutub nukuks ja vastne laguneb täielikult orgaanilisteks ühenditeks, millest ehitatakse uus organism.Täiskasvanud isend (imago) väljub nukust.
muna-vastne-nukk-imago
Kahepaiksed ja mõned putukad arenevad mittetäieliku transformatsiooniga
Siin ei ole nukku ja vastsete staadiumis toimub metamorfoos.
Muna-vastne-täiskasvanud
26. Inimese positsioon loomamaailma süsteemiga.
Bioloogia. Üldine bioloogia. 10. klass. Algtase Sivoglazov Vladislav Ivanovitš
16. Ainevahetus ja energia muundamine. Energiavahetus
Pea meeles!
Mis on ainevahetus?
Millisest kahest omavahel seotud protsessist see koosneb?
Kus inimkehas toimub enamiku toidust saadavate orgaaniliste ainete lagunemine?
Ainevahetus ja energia. Iga organismi elu peamine tingimus on ainete ja energia vahetus keskkonnaga. Igas rakus toimuvad pidevalt keerulised protsessid, mis on suunatud raku enda ja organismi kui terviku normaalse funktsioneerimise säilitamisele ja tagamisele. Sünteesitakse kompleksseid kõrgmolekulaarseid ühendeid: aminohapetest moodustuvad valgud, lihtsuhkrutest polüsahhariidid ja nukleotiididest nukleiinhapped. Rakud jagunevad ja moodustavad uusi organelle, erinevaid aineid transporditakse aktiivselt rakust ja rakku. Elektriimpulsid kanduvad edasi mööda närvikiude, lihased tõmbuvad kokku, püsib püsiv kehatemperatuur – see kõik, nagu ka paljud teised organismis toimuvad protsessid, nõuavad energiat. See energia tekib orgaaniliste ainete lagunemisel. Kõrgmolekulaarsete ühendite lõhustamisreaktsioonide kogum, millega kaasneb energia vabanemine ja salvestamine, kutsus energia metabolism või dissimilatsioon . Energia salvestatakse peamiselt universaalse energiamahuka ühendi - ATP kujul.
Adenosiintrifosforhape (ATP) on nukleotiid, mis koosneb lämmastikalusest (adeniinist), riboossuhkrust ja kolmest fosforhappejäägist (joonis 53). ATP on raku peamine energiamolekul, omamoodi energiaakumulaator. Kõigi protsessidega elusorganismides, mis nõuavad energiakulu, kaasneb ATP molekuli muundumine ADP-ks (adenosiindifosforhappeks). Fosforhappe jäägi elimineerimisel vabaneb suur hulk energiat - 40 kJ/mol. ATP molekulis on kaks sellist suure energiaga (nn kõrge energiaga) sidet. ATP struktuuri taastamine ADP-st ja fosforhappest toimub mitokondrites ja sellega kaasneb energia neeldumine.
Orgaaniliste ainete varu, mida keha kasutab energia saamiseks, tuleb pidevalt täiendada kas toiduga, nagu see juhtub loomadel, või sünteesi teel anorgaanilistest ainetest (taimedest). Kõikide elusorganismides toimuvate biosünteetiliste protsesside kogum, helistas plastivahetus või assimilatsioon . Plastivahetusega kaasneb alati energia neeldumine. Plastilise ainevahetuse peamised protsessid on valkude biosüntees (§ 13) ja fotosüntees (§ 17).
Riis. 53. ATP molekuli struktuur (märk "~" tähistab suure energiaga sidet)
Niisiis, energiavahetuse käigus lagundatakse orgaanilisi ühendeid ja salvestatakse energiat ning plastivahetuse käigus kulub energiat ja sünteesitakse orgaanilisi aineid. Energia ja plastilise vahetuse reaktsioonid on lahutamatult seotud, moodustades ühiselt ühe protsessi - ainevahetus ja energia , või ainevahetus . Ainevahetus toimub pidevalt kõigis rakkudes, kudedes ja elundites, säilitades keha sisekeskkonna püsivuse - homöostaas.
Energiavahetus. Enamik meie planeedi organisme vajab toimimiseks hapnikku. Selliseid organisme nimetatakse aeroobne. Energia metabolism aeroobides toimub kolmes etapis: ettevalmistav, hapnikuvaba ja hapnikuvaba. Hapniku juuresolekul oksüdeeruvad orgaanilised ained hingamisel täielikult süsihappegaasiks ja veeks, mille tulemusena salvestub suur hulk energiat.
Anaeroobsed organismid suudab ilma hapnikuta ellu jääda. Mõnele neist on hapnik üldiselt hävitav, mistõttu nad elavad seal, kus hapnikku üldse pole, näiteks teetanuse tekitaja. Teised, nn fakultatiivsed anaeroobid, võivad eksisteerida nii ilma hapnikuta kui ka selle juuresolekul. Energia metabolism anaeroobsetes organismides toimub kahes etapis: ettevalmistav ja hapnikuvaba, mistõttu orgaanilised ained ei oksüdeeru täielikult ja energiat talletatakse palju vähem.
Vaatleme kolme energiavahetuse etappi (joonis 54).
Ettevalmistav etapp. See etapp toimub seedetraktis ja rakkude lüsosoomides. Siin lagunevad kõrgmolekulaarsed ühendid seedeensüümide mõjul lihtsamateks madalamolekulaarseteks ühenditeks: valgud - aminohapeteks, polüsahhariidid - monosahhariidideks, rasvad - glütserooliks ja rasvhapeteks. Nende reaktsioonide käigus vabanevat energiat ei salvestata, vaid see hajub soojusena. Ettevalmistavas etapis moodustunud madala molekulmassiga aineid saab organism kasutada oma orgaaniliste ühendite sünteesimiseks, st siseneda plastilisse ainevahetusse või energia salvestamiseks edasi lagundada.

Riis. 54. Energiavahetuse etapid
Hapnikuvaba etapp. Teine etapp toimub rakkude tsütoplasmas, kus toimub edasine lihtsate orgaaniliste ainete lagunemine. Keha ei kasuta esimeses etapis tekkinud aminohappeid järgmistes dissimilatsioonietappides, sest vajab neid materjalina omaenda valgumolekulide sünteesiks. Seetõttu kasutatakse valke energia saamiseks väga harva, enamasti alles siis, kui ülejäänud varud (süsivesikud ja rasvad) on juba ammendatud. Tavaliselt on rakus kõige kättesaadavam energiaallikas glükoos.
Glükoosi hapnikuvaba lagunemise keerulist mitmeastmelist protsessi energia metabolismi teises etapis nimetatakse glükolüüs(kreeka keelest glükoosid- magus ja lüüsimine– poolitamine).
Glükolüüsi tulemusena laguneb glükoos lihtsamateks orgaanilisteks ühenditeks (glükoos C 6 H 12 O 6 - püroviinamarihape C 3 H 4 O 3). See vabastab energiat, millest 60% hajub soojusena ja 40% kasutatakse ATP sünteesiks. Ühe glükoosi molekuli lagundamisel moodustub kaks ATP molekuli ja kaks püroviinamarihappe molekuli. Seega hakkab keha dissimilatsiooni teises etapis energiat talletama.
Püruviinhappe edasine saatus sõltub hapniku olemasolust rakus. Kui hapnik on olemas, siis siseneb püroviinamarihape mitokondritesse, kus see oksüdeerub täielikult CO 2 -ks ja H 2 O-ks ning toimub energiavahetuse kolmas, hapniku staadium (vt allpool).
Hapniku puudumisel toimub nn anaeroobne hingamine, mida sageli nimetatakse kääritamine. Pärmirakkudes muundatakse alkoholkäärimise käigus püroviinamarihape (PVA) etüülalkoholiks (PVA? Etüülalkohol + CO 2).
Piimhappekäärimise käigus tekib PVC-st piimhape. See protsess võib toimuda mitte ainult piimhappebakterites. Raske füüsilise töö käigus tekib inimese lihaskoe rakkudes hapnikupuudus, mille tagajärjel tekib piimhape, mille kuhjumine põhjustab väsimustunnet, valu ja vahel isegi krampe.
Hapniku staadium. Kolmandas etapis oksüdeeritakse glükoosi hapnikuvaba lagunemise käigus tekkinud saadused süsinikdioksiidiks ja veeks. Nii vabaneb suur hulk energiat, millest märkimisväärne osa kulub ATP sünteesiks. See protsess toimub mitokondrites ja seda nimetatakse rakuhingamine. Rakulise hingamise käigus vabaneb kahe PVC-molekuli oksüdeerumisel kehas talletatud energia 36 ATP-molekuli kujul.
Niisiis moodustub energia metabolismi protsessis ühe glükoosimolekuli täieliku oksüdeerumisega süsinikdioksiidiks ja veeks 38 ATP molekuli (2 molekuli glükolüüsi protsessis ja 36 raku hingamise protsessis mitokondrites):
C6H12O6 + 6O2 + 38ADP + 38F 6CO 2? 6H20 + 38ATP.
Anaeroobsetes tingimustes on energia metabolismi efektiivsus palju madalam – ainult 2 ATP molekuli. Käärimisproduktid (etüülalkohol, piimhape, võihape) säilitavad oma keemilistes sidemetes endiselt palju energiat, st hapniku dissimilatsioonitee on energeetiliselt soodsam. Kuid ajalooliselt on kääritamine iidsem protsess. Seda oleks võinud läbi viia ka siis, kui iidse Maa atmosfääris polnud vaba hapnikku.
Vaadake üle küsimused ja ülesanded
1. Mis on dissimilatsioon? Loetlege selle etapid.
2. Milline on ATP roll rakkude metabolismis?
3. Millised rakustruktuurid teostavad ATP sünteesi?
4. Selgitage energia metabolismi rakus, kasutades näitena glükoosi lagunemist.
5. Joonistage dissimilatsiooniprotsessi skemaatiline diagramm, võttes ühele diagrammile kokku kõik selle lõigu tekstis mainitud võimalikud variandid (sh kääritamine).
6. Sõnade "dissimilatsioon" ja "assimilatsioon" sünonüümid on terminid "katabolism" ja "anabolism". Selgitage nende mõistete päritolu.
mõtle! Tee seda!
1. Selgitage, miks liigne söömine põhjustab rasvumist.
2. Miks ei saa energiavahetus eksisteerida ilma plastivahetuseta?
3. Miks arvate, et pärast rasket füüsilist tööd on lihasvalu kiireks leevendamiseks soovitatav minna sooja vanni?
Töö arvutiga
Vaadake elektroonilist taotlust. Tutvuge materjaliga ja täitke ülesanded.
Raamatust Teeninduskoer [Teenistuskoerte aretusspetsialistide koolituse juhend] autor Krušinski Leonid Viktorovitš3. Ainevahetus kui elu alus “Elu on valgukehade eksisteerimise vorm,” kirjutas F. Engels. Seetõttu võime öelda, et valk on elu kandja. Valk on keeruline aine, mis koosneb paljudest elementidest, sealhulgas lämmastiku olemasolust.
Raamatust Ajastu anatoomia ja füsioloogia autor Antonova Olga AleksandrovnaTeema 10. AINEVAHETUSE JA ENERGIA VANUSED 10.1. Ainevahetusprotsesside tunnused Ainevahetus ja energia on organismi elutähtsate protsesside aluseks. Inimese kehas, tema organites, kudedes, rakkudes toimub pidev sünteesiprotsess, s.o.
Raamatust Bioloogia [Täielik teatmik ühtseks riigieksamiks valmistumiseks] autor Lerner Georgi Isaakovitš Raamatust Stopp, kes juhib? [Inimeste ja teiste loomade käitumise bioloogia] autor Žukov. Dmitri Anatoljevitš Raamatust Lood bioenergiast autor Skulatšov Vladimir Petrovitš Raamatust Bioloogia. Üldine bioloogia. 11. klass. Põhitase autor Sivoglazov Vladislav Ivanovitš Raamatust Inimese pärilikkuse saladused autor Afonkin Sergei Jurjevitš Raamatust Antropoloogia ja bioloogia mõisted autor Kurtšanov Nikolai Anatolievitš Raamatust Bioloogiline keemia autor Lelevitš Vladimir Valerjanovitš Autori raamatustSÜSIVESIKUTE AINEVAHETUS Tuleb veel kord rõhutada, et kehas toimuvad protsessid kujutavad endast ühtset tervikut ning ainult esitluse mugavuse ja tajumise hõlbustamiseks käsitletakse neid õpikutes ja käsiraamatutes eraldi peatükkidena. See kehtib ka jagunemise kohta
Autori raamatustPeatükk 2. Mis on energia metabolism? Kuidas rakk energiat vastu võtab ja kasutab Elamiseks on vaja tööd teha. See igapäevane tõde on üsna kohaldatav igale elusolendile. Kõik organismid: alates üherakulistest mikroobidest kuni kõrgemate loomade ja inimesteni - tegutsevad pidevalt
Autori raamatust25. Toiduühendused. Aine ja energia tsükkel ökosüsteemides Pidage meeles! Millised olulised komponendid sisalduvad igas ökosüsteemis? Elusorganismid on pidevas vastasmõjus üksteise ja keskkonnateguritega, moodustades jätkusuutliku
Autori raamatustAinevahetus Meie haigused on endiselt samad, mis tuhandeid aastaid tagasi, kuid arstid on neile leidnud kallimad nimed. Rahvatarkus - Kõrge kolesteroolitase võib olla päritav - Varajane suremus ja kolesterooli kasutamise eest vastutavad geenid - Kas see on päritav
Autori raamatust2.3. Ainevahetus ja energia Kogu elusorganismides toimuvate keemiliste reaktsioonide kogumit nimetatakse ainevahetuseks ehk ainevahetuseks. Nende reaktsioonide tulemusena muundub keemilistes sidemetes salvestunud energia muudeks vormideks, st ainevahetus toimub alati
Autori raamatustPeatükk 10. Energia ainevahetus. Bioloogiline oksüdatsioon Elusorganismid on termodünaamika seisukohalt avatud süsteemid. Süsteemi ja keskkonna vahel on võimalik energiavahetus, mis toimub kooskõlas termodünaamika seadustega. Iga orgaaniline
Kõik elusorganismid vahetavad aineid väliskeskkonnaga. Rakkudes toimuvad pidevalt biosünteesi protsessid. Tänu ensüümidele tekivad lihtsatest ainetest kompleksühendid: aminohapetest sünteesitakse valgud, monosahhariididest komplekssüsivesikud ja lämmastikalustest nukleiinhapped. Suhteliselt lihtsate ainete keemilisel muundamisel tekivad mitmesugused rasvad ja õlid. Kitiin on lülijalgsete väliskate, moodustades kitiini - kompleksse polüsahhariidi (lk 7), lindudel ja imetajatel on väliskatteks sarvjas aine, mille aluseks on valkkeratiin. Lõppkokkuvõttes määrab sünteesitud suurte orgaaniliste molekulide koostise genotüüp. Sünteesitud aineid kasutatakse kasvu ajal rakkude ja nende organellide ehitamiseks ning kulunud või hävinud molekulide asendamiseks. Eranditult toimuvad kõik biosünteesi vastasmõjud energia neeldumisega.
Plastivahetus
Plastiline metabolism, mida muidu nimetatakse biosünteesiks või anabolismiks, toimub see vahetus ainult rakus. Plastilist ainevahetust on kolme tüüpi: fotosüntees, kemosüntees ja valkude biosüntees. Fotosünteesi kasutavad taimed ja ainult mõned bakterid (tsüanobakterid). Selliseid organisme nimetatakse autotroofideks. Kemosünteesi kasutavad teatud bakterid, sealhulgas anaeroobsed. Selliseid organisme nimetatakse kemotroofideks. Loomad ja seened liigitatakse heterotroofseteks olenditeks.
Fotosüntees
Fotosünteesi protsess toimub reaktsiooni kaudu, mis hõlmab glükoosi ja hapniku moodustumist süsinikdioksiidist ja veest. Fotosünteesil on kaks faasi, hele ja tume. Valgusfaasis toimub fotosünteesi protsess kloroplasti graanas ja pimedas faasis kloroplasti stroomas. (vt 7. lisa). Ilma päikeseenergiata poleks fotosünteesil tähtsust, seega on see oluline tegur. Selle protsessi käigus moodustub kuuest hapnikumolekulist ja ühest glükoosi molekulist kuus süsinikdioksiidi ja vee molekuli. Fotosünteesi protsess toimub kloroplastides, klorofülli leidub organellides, tänu millele toimub süntees.
6СО2 + 6Н2О → С6Н12О6 + 6О2
Kemosüntees
Kemosüntees on iseloomulik sellistele bakteritele nagu väävli-, nitrifitseerimis- ja rauabakterid. Bakterid kasutavad ainete oksüdatsiooni käigus omandatud energiat süsinikdioksiidi redutseerimiseks orgaanilisteks ühenditeks. (vt 8. lisa) Väävlibakterid oksüdeerivad selliseid aineid nagu vesiniksulfiid, nitrifitseerivad bakterid oksüdeerivad ammoniaaki ja rauabakterid raudoksiidi.
Valkude biosüntees
Plastiline metabolism on valkude süntees raku poolt. Vahetusel on kaks peamist protsessi: transkriptsioon ja tõlkimine.
Transkriptsioon- See on messenger-RNA sünteesimise protsess, kasutades DNA-d vastavalt komplementaarsuse põhimõttele. (vt 9. lisa)
Transkriptsioonil on kolm etappi:
Esmane transkriptsiooni moodustamine
Töötlemine
Splaissimine
Saade- valgu struktuuri kohta teabe edastamine messenger-RNA-lt sünteesitud polüpeptiidile. (vt 10. lisa) See protsess viiakse läbi tsütoplasmas ribosoomil. Ülekanne toimub neljas etapis. Esimesel etapil aktiveeritakse aminohapped spetsiaalse ensüümi - aminoatsüül-T-RNA süntetaasiga. See protsess kasutab energiat ATP kujul. Seejärel moodustub minoatsüüladenülaat. Sellele järgneb aktiveeritud aminohappe külgnemise protsess ülekande-RNA-ga ja AMP vabaneb. Lisaks seostub moodustunud kompleks kolmandas etapis ribosoomiga. Seejärel lülitatakse aminohapped valgu struktuuri kindlas järjestuses, mille järel vabaneb ülekande-RNA.
Energiavahetus
Energia metabolismi nimetatakse ka katabolismiks. Plastik ja energiaainevahetus on omavahel väga seotud, sest plastilise ainevahetuse (anabolismi) läbiviimiseks on vaja energiat, mida rakk saab katabolismi teel. Seda protsessi kasutades sünteesib rakk vajalikud nukleiinhapped, valgud, süsivesikud jne. Energiaainevahetus on protsess, mille käigus lagundatakse keerulise ehitusega ained lihtsamateks või oksüdeeritakse, mille tõttu organism omandab eksisteerimiseks vajaliku energia. Energia metabolismis on kolm etappi:
Ettevalmistav etapp
Anaeroobne staadium - glükolüüs (ilma hapnikuta)
Aeroobne staadium - rakuline hingamine (hapniku osalusel)
Ettevalmistav etapp
Selles etapis muudetakse polümeerid monomeerideks, st sellised ühendid nagu valgud, süsivesikud ja lipoidid lagundatakse lihtsamateks. See protsess toimub väljaspool rakku, seedesüsteemi organites. Selles energiavahetuse etapis ei ole hapnikku vaja. Reaktsioonide tulemusena laguneb valk aminohapeteks, liitsüsivesikud lihtmonosahhariidideks ning lipiidid glütserooliks ja kõrgemateks hapeteks. See etapp esineb ka raku lüsosoomides.
Anaeroobne etapp
Seda etappi nimetatakse ka fermentatsiooniks või glükolüüsiks. Ettevalmistavas etapis moodustunud ained - glükoos, aminohapped jne - läbivad järgneva ensümaatilise lagunemise ilma hapniku osaluseta. Süsivesikud on peamiselt kääritatud. Selles katabolismi etapis kasutatavate keemiliste reaktsioonide käigus tekivad alkoholid, süsinikdioksiid, atsetoon, orgaanilised happed ning mõnel juhul vesinik ja muud ained. Glükolüüs on glükoosi lagunemine anaeroobsetes tingimustes püroviinamarihappeks (PVA), seejärel piim-, äädik-, võihappeks või etüülalkoholiks, mis toimub raku tsütoplasmas. Hapnikuvaba lõhustumise käigus hajub osa vabanenud energiast soojuse kujul ja osa salvestub ATP molekulidesse. Tavaline reaktsioon loomade ja seente rakkudes on püroviinamarihappe vabanemine.
Põhiline keemiline reaktsioon selles etapis näeb välja järgmine:
C6H12O6 = 2C3H4O3 + (4H) + 2ATP
Selle protsessi tulemusena moodustub kaks ATP molekuli.
Aeroobne etapp
See etapp toimub mitokondrites. (vt lisa 11) Selles etapis ained oksüdeeritakse, mille tõttu vabaneb teatud kogus energiat. Hapnik osaleb samas protsessis. Hapnikku transporditakse hemoglobiini sisaldavate punaste vereliblede abil. Eelmistes etappides saadud ained lagundatakse raku poolt kõige lihtsamateks ehk süsihappegaasiks ja veeks. Lüsosoomides sisalduvad ensüümid oksüdeerivad rakus orgaanilisi ühendeid. ADP - adenosiindifosfaat - aine, mis on rakuhingamise tõttu vajalik ka energia tootmiseks. Põhiline keemiline reaktsioon selles etapis näeb välja järgmine:
2C3H6O3 + 6O2 + 36H3PO4 + 36ADP = 6CO2 + 42H2O + 36ATP
Selle protsessi tulemusena moodustub 36 ATP molekuli.
Sellest võrrandist näete, et selles etapis vabaneb märkimisväärne kogus energiat. Lisaks võib selles etapis toimuda püroviinamarihappe täieliku oksüdatsiooni reaktsioon, mille tulemusena vabaneb ka energia, kuid väiksemates kogustes.
Järelikult suudab rakk ühe glükoosimolekuli täielikul lagunemisel sünteesida 38 ATP molekuli (2 molekuli glükolüüsi ajal ja 36 molekuli aeroobses etapis). (vt lisa 12)
Aeroobse hingamise üldvõrrandi saab kirjutada järgmiselt:
C6H1206 + 602 + 38ADP + 38H3P04 > 6C02 + 6H20 + 38ATP.
Järeldus
Rakk on kõrgelt organiseeritud eluüksus. Ainete ja energia imendumine, muundumine, säilitamine ja kasutamine toimub rakkude kaudu. Just rakus toimuvad sellised protsessid nagu hingamine, fermentatsioon, fotosüntees ja geneetilise materjali dubleerimine. Ja sellised protsessid toimuvad nii lihtsa ehitusega organismides (üherakulised) kui ka keeruka ehitusega organismides (mitmerakulised). Kõigi organismide eluiga sõltub nende rakkudest.
Rakendus
Lisa 1
2. lisa

3. lisa

4. lisa

5. lisa

6. lisa

7. lisa

8. lisa

9. lisa

Ainete ja energia ainevahetus (ainevahetus) toimub keha kõigil tasanditel: raku-, koe- ja organismitasanditel. See tagab keha sisekeskkonna – homöostaasi – püsivuse pidevalt muutuvates eksistentsitingimustes. Rakus toimuvad samaaegselt kaks protsessi – plastiline ainevahetus (anabolism või assimilatsioon) ja energia metabolism (fatabolism või dissimilatsioon).
Plastiline ainevahetus on biosünteesireaktsioonide kogum ehk lihtsatest molekulidest keeruliste molekulide loomine. Rakk sünteesib pidevalt aminohapetest valke, glütseroolist ja rasvhapetest rasvu, monosahhariididest süsivesikuid, lämmastikalustest nukleotiide ja suhkruid. Need reaktsioonid nõuavad energiat. Kasutatav energia vabaneb energiavahetuse teel. Energia metabolism on reaktsioonide kogum, mis lagundab keerulised orgaanilised ühendid lihtsamateks molekulideks. Osa sel juhul vabanevast energiast läheb energiasidemetega rikaste ATP (adenosiintrifosforhappe) molekulide sünteesiks. Orgaaniliste ainete lagunemine toimub tsütoplasmas ja mitokondrites hapniku osalusel. Assimilatsiooni- ja dissimilatsioonireaktsioonid on üksteise ja väliskeskkonnaga tihedalt seotud. Keha saab toitaineid väliskeskkonnast. Jäätmed satuvad väliskeskkonda.
Ensüümid (ensüümid) on spetsiifilised valgud, bioloogilised katalüsaatorid, mis kiirendavad metaboolseid reaktsioone rakus. Kõik protsessid elusorganismis toimuvad otseselt või kaudselt ensüümide osalusel. Ensüüm katalüüsib ainult ühte reaktsiooni või toimib ainult ühte tüüpi sidemetele. See tagab kõigi rakus või kehas toimuvate elutähtsate protsesside (hingamine, seedimine, fotosüntees jne) peenregulatsiooni. Iga ensüümi molekulis on koht, mis loob kontakti ensüümi molekulide ja konkreetse aine (substraadi) vahel. Ensüümi aktiivne keskus on funktsionaalne rühm (näiteks OH - seriinirühm) või eraldi aminohape.
Ensümaatiliste reaktsioonide kiirus sõltub paljudest teguritest: temperatuur, rõhk, keskkonna happesus, inhibiitorite olemasolu jne.
Energia metabolismi etapid:
- Ettevalmistav- esineb rakkude tsütoplasmas. Ensüümide toimel lagunevad polüsahhariidid monosahhariidideks (glükoos, fruktoos jne), rasvad glütserooliks ja rasvhapeteks, valgud aminohapeteks ning nukleiinhapped nukleotiidideks. See vabastab väikese koguse energiat, mis hajub soojusena.
- Hapnikuvaba(anaeroobne hingamine või glükolüüs) - glükoosi mitmeastmeline lagunemine ilma hapniku osaluseta. Seda nimetatakse kääritamiseks. Lihastes laguneb glükoosimolekul anaeroobse hingamise tulemusena kaheks lüüroviinahappe (C 3 H 4 O 3) molekuliks, mis seejärel taandatakse piimhappeks (C 3 H 6 O 3). Fosforhape ja ADP osalevad glükoosi lagundamisel.
Selle etapi üldvõrrand: C 6 H 12 O 6 + 2H 3 PO 4 + 2ADP -> 2C 3 H 6 O 3 + 2ATP + 2H 2 O
Pärmseentes muundatakse glükoosi molekul ilma hapniku osaluseta etüülalkoholiks ja süsinikdioksiidiks (alkohoolne käärimine). Teistes mikroorganismides võib glükolüüsi tulemusena tekkida atsetoon, äädikhape jne. Ühe glükoosimolekuli lagunemisel tekib kaks ATP molekuli, mille sidemetes salvestub 40% energiast, ülejäänud energia hajub soojuse vorm.
- Hapniku hingamine- aeroobse hingamise ehk hapniku lõhustumise staadium, mis toimub mitokondrite sisemembraani voltidel - cristae. Selles etapis lagundatakse eelmise etapi ained lõplikeks laguproduktideks – veeks ja süsihappegaasiks. Kahe piimhappemolekuli lagunemise tulemusena moodustub 36 ATP molekuli. Hapniku lagunemise normaalse kulgemise peamine tingimus on mitokondriaalsete membraanide terviklikkus. Hapnikuhingamine on peamine samm rakkude hapnikuga varustamisel. See on 20 korda tõhusam kui hapnikuvaba etapp.
Hapniku jagunemise üldvõrrand on: 2C 3 H 6 0 3 + 60 2 + 36H 3 PO 4 + 36ADP -> 6CO 2 + 38H 2 O + 36ATP
Energia saamise meetodi järgi jagunevad kõik organismid kahte rühma - autotroofsed ja heterotroofsed.
Energia metabolism taimede, seente ja loomade aeroobsetes rakkudes toimub samamoodi. See näitab nende suhet. Mitokondrite arv koerakkudes on erinev, see sõltub rakkude funktsionaalsest aktiivsusest. Näiteks lihasrakkudes on palju mitokondreid.
Rasvade lagunemine glütserooliks ja rasvhapeteks toimub ensüümide – lipaaside – abil. Valgud lagundatakse esmalt oligopeptiidideks ja seejärel aminohapeteks.
Ensüümid (ladina keelest "fermentum" - käärimine, juuretis), ensüümid, spetsiifilised valgud, mis suurendavad keemiliste reaktsioonide kiirust kõigi elusorganismide rakkudes. Keemilise olemuse järgi - valgud, millel on optimaalne aktiivsus teatud pH juures, vajalike koensüümide ja kofaktorite olemasolu ning inhibiitorite puudumine. Ensüüme nimetatakse ka biokatalüsaatoriteks analoogselt keemia katalüsaatoritega. Iga ensüümi tüüp katalüüsib teatud ainete (substraatide) muundumist, mõnikord ainult ühe aine ühes suunas. Seetõttu viib rakkudes läbi arvukalt biokeemilisi reaktsioone tohutu hulk erinevaid ensüüme. Need jagunevad 6 klassi: oksidoreduktaasid, transferaasid, hüdrolaasid, lüaasid, isomeraasid ja ligaasid. Paljud ensüümid on eraldatud elusrakkudest ja saadud kristallilisel kujul (esimest korda 1926. aastal).
Ensüümide roll organismis
Ensüümid osalevad kõigis ainevahetusprotsessides ja geneetilise informatsiooni rakendamises. Toitainete seedimine ja assimilatsioon, valkude, nukleiinhapete, rasvade, süsivesikute ja muude ühendite süntees ja lagunemine kõigi organismide rakkudes ja kudedes – kõik need protsessid on võimatud ilma ensüümide osaluseta. Igasugune elusorganismi funktsioonide - hingamine, lihaste kokkutõmbumine, neuropsüühiline aktiivsus, paljunemine jne - avaldumine on tagatud ensüümide tegevusega. Teatud funktsioone täitvate rakkude individuaalsed omadused on suuresti määratud ainulaadse ensüümide komplektiga, mille tootmine on geneetiliselt programmeeritud. Isegi ühe ensüümi puudumine või selle mis tahes defekt võib põhjustada kehale tõsiseid negatiivseid tagajärgi.
Ensüümide katalüütilised omadused
Ensüümid on kõigist teadaolevatest katalüsaatoritest kõige aktiivsemad. Enamik reaktsioone rakus kulgeb miljoneid ja miljardeid kordi kiiremini kui siis, kui need toimuksid ensüümide puudumisel. Seega on üks katalaasi ensüümi molekul võimeline teisendama sekundiga kuni 10 tuhat erinevate ühendite oksüdatsiooni käigus tekkinud rakkudele mürgist vesinikperoksiidi molekuli veeks ja hapnikuks. Ensüümide katalüütilised omadused tulenevad nende võimest oluliselt vähendada reageerivate ühendite aktivatsioonienergiat, st ensüümide juuresolekul kulub antud reaktsiooni “käivitamiseks” vähem energiat.
Ensüümide avastamise ajalugu
Ensüümide osalusel toimuvad protsessid on inimesele teada iidsetest aegadest, sest leiva, juustu, veini ja äädika valmistamine põhineb ensümaatilistel protsessidel. Kuid alles 1833. aastal eraldati idanevatest odrateradest esimest korda toimeaine, mis muutis tärklise suhkruks ja nimetati diastaasiks (nüüd nimetatakse seda ensüümi amülaasiks). 19. sajandi lõpus. On tõestatud, et pärmirakkude jahvatamisel saadud mahl sisaldab keerulist ensüümide segu, mis tagab alkohoolse käärimise protsessi. Sellest ajast algas intensiivne ensüümide – nende struktuuri ja toimemehhanismi – uurimine. Kuna kääritamise uurimisel ilmnes biokatalüüsi roll, siis alates 19. sajandist on selle protsessiga seostatud kahte väljakujunenud protsessi. nimetused on "ensüüm" (tõlkes kreeka keelest "pärmist") ja "ensüüm". Tõsi, viimast sünonüümi kasutatakse ainult venekeelses kirjanduses, kuigi ensüümide ja nende osalusega protsesside uurimisega seotud teaduslikku suunda nimetatakse traditsiooniliselt ensümoloogiaks. 20. sajandi esimesel poolel. Tehti kindlaks, et keemilise olemuselt on ensüümid valgud ja sajandi teisel poolel määrati juba paljudele sadadele ensüümidele aminohappejääkide järjestus ning pandi paika ruumiline struktuur. 1969. aastal viidi esmakordselt läbi ensüümi ribonukleaasi keemiline süntees. Ensüümide toimemehhanismi mõistmisel on tehtud tohutuid edusamme.
Ensüümide asukoht kehas
Rakus asuvad mõned ensüümid tsütoplasmas, kuid enamasti on ensüümid seotud teatud rakustruktuuridega, kus nad oma toimet avaldavad. Näiteks tuumas on ensüümid, mis vastutavad replikatsiooni - DNA sünteesi (DNA polümeraas) ja selle transkriptsiooni - RNA (RNA polümeraasi) moodustumise eest. Mitokondrid sisaldavad ensüüme, mis vastutavad energia salvestamise eest; lüsosoomid sisaldavad enamikku hüdrolüütilistest ensüümidest, mis on seotud nukleiinhapete ja valkude lagunemisega.
Ensüümi toimetingimused
Kõik ensüümidega seotud reaktsioonid toimuvad peamiselt neutraalses, kergelt aluselises või kergelt happelises keskkonnas. Iga üksiku ensüümi maksimaalne aktiivsus ilmneb aga rangelt määratletud pH väärtuste juures. Enamiku ensüümide toimeks soojaverelistel loomadel on soodsaim temperatuur 37-40oC. Taimedel temperatuuril alla 0o C ei lakka ensüümide toime täielikult, kuigi taimede elutegevus väheneb järsult. Ensümaatilised protsessid ei saa reeglina toimuda temperatuuril üle 70 ° C, kuna ensüümid, nagu kõik valgud, alluvad termilisele denaturatsioonile (struktuuride hävitamisele).
Ensüümide suurused ja struktuur
Ensüümide, nagu kõigi teiste valkude, molekulmass jääb vahemikku 10 tuhat kuni 1 miljon (aga võib olla ka rohkem). Need võivad koosneda ühest või mitmest polüpeptiidahelast ja neid võivad esindada kompleksvalgud. Viimane sisaldab koos valgukomponendiga (apoensüüm) madalmolekulaarseid ühendeid - koensüüme (kofaktoreid, koensüüme), sealhulgas metalliioone, nukleotiide, vitamiine ja nende derivaate. Mõned ensüümid moodustuvad inaktiivsete prekursorite (proensüümide) kujul ja muutuvad aktiivseks pärast teatud muutusi molekuli struktuuris, näiteks pärast väikese fragmendi eraldamist sellest. Nende hulka kuuluvad seedeensüümid trüpsiin ja kümotrüpsiin, mida pankrease rakud sünteesivad mitteaktiivsete lähteainetena (trüpsinogeen ja kümotrüpsinogeen) ja muutuvad aktiivseks peensooles pankrease mahla osana. Paljud ensüümid moodustavad nn ensüümikomplekse. Sellised kompleksid on näiteks põimitud rakkude membraanidesse või raku organellidesse ja osalevad ainete transpordis.
Transformatsioonis olev aine (substraat) seondub ensüümi kindla osaga, aktiivse tsentriga, mille moodustavad aminohapete külgahelad, mis paiknevad sageli polüpeptiidahela osades, mis on üksteisest märkimisväärselt kaugel. Näiteks kümotrüpsiini molekuli aktiivse tsentri moodustavad histidiini jäägid, mis asuvad polüpeptiidahelas positsioonis 57, seriin positsioonis 195 ja asparagiinhape positsioonis 102 (kokku on kümotrüpsiini molekulis 245 aminohapet). Seega annab polüpeptiidahela kompleksne paigutus valgu molekulis - ensüümis võimaluse mitmel aminohappe kõrvalahelal tekkida rangelt määratletud kohas ja teatud kaugusel üksteisest. Aktiivse tsentri osaks on ka koensüümid (valguline osa ja mittevalguline komponent eraldi ei oma ensümaatilist aktiivsust ja omandavad ensüümi omadused ainult koos kombineerituna).
Ensüümidega seotud protsessid
Enamikku ensüüme iseloomustab toime kõrge spetsiifilisus (selektiivsus), kui iga reagendi (substraadi) muundamine reaktsioonisaaduseks viiakse läbi spetsiaalse ensüümi abil. Sel juhul võib ensüümi toime olla rangelt piiratud ühe substraadiga. Näiteks ensüüm ureaas, mis osaleb karbamiidi lagunemisel ammoniaagiks ja süsihappegaasiks, ei reageeri sarnase struktuuriga metüüluureaga. Paljud ensüümid toimivad mitmele struktuurilt sarnasele ühendile või ühte tüüpi keemilistele sidemetele (näiteks fosfodiestersideme lõhustavad ensüümfosfataasid). Ensüüm toimib ensüümi-substraadi kompleksi moodustamise kaudu, mis seejärel laguneb, moodustades ensümaatilise reaktsiooni produktid ja vabastades ensüümi. Ensüüm-substraadi kompleksi moodustumise tulemusena muudab substraat oma konfiguratsiooni; sel juhul muundatud ensüüm-keemiline side nõrgeneb ja reaktsioon kulgeb väiksema algse energiakuluga ja seetõttu palju suurema kiirusega. Ensümaatilise reaktsiooni kiirust mõõdetakse ajaühikus muundatud substraadi koguse või moodustunud produkti kogusega. Paljud ensümaatilised reaktsioonid, olenevalt substraadi ja reaktsioonisaaduse kontsentratsioonist keskkonnas, võivad toimuda nii edasi- kui ka vastupidises suunas (substraadi liig nihutab reaktsiooni saaduse moodustumise suunas, samas kui substraadi kogunemine toimub liigselt Viimasel juhul toimub substraadi süntees). See tähendab, et ensümaatilised reaktsioonid võivad olla pöörduvad. Näiteks süsihappeanhüdraas veres muudab kudedest tuleva süsihappegaasi süsihappeks (H2CO3), kopsudes vastupidi, katalüüsib süsihappe muutumist veeks ja süsihappegaasiks, mis eemaldatakse väljahingamisel. Siiski tuleb meeles pidada, et ensüümid, nagu ka teised katalüsaatorid, ei saa nihutada keemilise reaktsiooni termodünaamilist tasakaalu, vaid ainult kiirendavad oluliselt selle tasakaalu saavutamist.
Ensüümide nimede nomenklatuur
Ensüümi alusena nimetamisel võtke substraadi nimi ja lisage järelliide “aza”. Nii tekkisid eelkõige proteinaasid - ensüümid, mis lagundavad valke (valke), lipaase (lagustab lipiide või rasvu) jne. Mõned ensüümid said erilised (triviaalsed) nimed, näiteks seedeensüümid - pepsiin, kümotrüpsiin ja trüpsiin. .
Keharakkudes toimub mitu tuhat erinevat metaboolset reaktsiooni ja seetõttu on ensüüme sama palju. Sellise mitmekesisuse süsteemi toomiseks võeti vastu rahvusvaheline leping ensüümide klassifitseerimise kohta. Selle süsteemi kohaselt jaotati kõik ensüümid, olenevalt nende katalüüsitavate reaktsioonide tüübist, kuueks põhiklassiks, millest igaüks sisaldab mitmeid alamklasse. Lisaks sai iga ensüüm neljakohalise koodinumbri (šifri) ja nime, mis näitab reaktsiooni, mida ensüüm katalüüsib. Erinevate liikide organismides sama reaktsiooni katalüüsivad ensüümid võivad oma valgustruktuurilt oluliselt erineda, kuid nomenklatuuris on neil ühine nimetus ja üks koodnumber.
Ensüümide tootmise häiretega seotud haigused
Mis tahes ensüümi aktiivsuse puudumine või vähenemine (sageli ülemäärane aktiivsus) inimestel põhjustab haiguste (ensümopaatiate) arengut või keha surma. Seega areneb laste pärilik haigus - galaktoseemia (viib vaimse alaarenguni) - galaktoosi kergesti seeditavaks glükoosiks muutmise eest vastutava ensüümi sünteesi rikkumise tagajärjel. Teise päriliku haiguse - fenüülketonuuria, millega kaasneb vaimse aktiivsuse häire, põhjuseks on maksarakkude võime sünteesida ensüümi, mis katalüüsib aminohappe fenüülalaniini türosiiniks muutumist. Paljude ensüümide aktiivsuse määramist veres, uriinis, tserebrospinaal-, seemne- ja teistes kehavedelikes kasutatakse mitmete haiguste diagnoosimiseks. Selle vereseerumi analüüsi abil on võimalik varajases staadiumis avastada müokardiinfarkti, viirushepatiiti, pankreatiiti, nefriiti ja muid haigusi.
Ensüümide kasutamine inimestel
Kuna ensüümid säilitavad oma omadused väljaspool keha, kasutatakse neid edukalt erinevates tööstusharudes. Näiteks papaia proteolüütiline ensüüm (papaya mahlast) - pruulimisel, liha pehmendamiseks; pepsiin - "valmis" teravilja tootmisel ja ravimina; trüpsiin - imikutoidutoodete tootmisel; renniin (vasika maost pärinev laap) - juustu valmistamisel. Katalaasi kasutatakse laialdaselt toiduainete- ja kummitööstuses, puuviljamahlade selitamiseks kasutatakse polüsahhariide lagundavaid tsellulaase ja pektidaase. Ensüümid on vajalikud valkude, nukleiinhapete ja polüsahhariidide struktuuri kindlakstegemisel, geenitehnoloogias jne. Ensüümide abil saadakse ravimeid ja kompleksseid keemilisi ühendeid.
On avastatud teatud ribonukleiinhapete (ribosüümide) vormide võime katalüüsida üksikuid reaktsioone, st toimida ensüümidena. Võib-olla toimisid orgaanilise maailma evolutsiooni ajal ribosüümid biokatalüsaatoritena, enne kui ensümaatiline funktsioon kanti üle selle ülesande täitmiseks paremini sobivatele valkudele.
 Kehade liikumine gravitatsiooni mõjul
Kehade liikumine gravitatsiooni mõjul Troobid ja stiilifiguurid
Troobid ja stiilifiguurid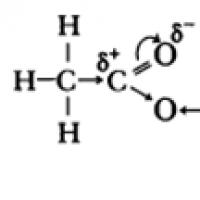 §12. Karboksüülhapped. Karboksüülhapped: füüsikalised omadused. Karboksüülhapete soolad Lihtsaim karboksüülhape on sipelghape
§12. Karboksüülhapped. Karboksüülhapped: füüsikalised omadused. Karboksüülhapete soolad Lihtsaim karboksüülhape on sipelghape Vestlus endise tüdruksõbraga
Vestlus endise tüdruksõbraga Miks sa unistad oma parimast sõbrast, endisest sõbrast või lapsepõlvesõbrast?
Miks sa unistad oma parimast sõbrast, endisest sõbrast või lapsepõlvesõbrast? Raseda naise unenägude tõlgendus unistab lapsest
Raseda naise unenägude tõlgendus unistab lapsest Miks unistad sädevoolust? Miks sa unistad sädemetest? Mida nad unenägude raamatutes kirjutavad
Miks unistad sädevoolust? Miks sa unistad sädemetest? Mida nad unenägude raamatutes kirjutavad